

Grégory Beaugrand, Université de Lille
Une des questions fondamentales en biologie est de comprendre comment la biodiversité s’organise sur notre planète. Certaines interrogations restent en effet sans réponse. Pourquoi y a-t-il plus d’espèces terrestres que marines, (environ six fois plus) ? Cette énigme est d’autant plus importante que la biodiversité marine est apparue bien avant celle des terres émergées de notre planète. Puisque plus de temps s’est écoulé depuis l’émergence de la vie dans les océans, la supériorité du nombre d’espèces terrestres par rapport aux espèces marines semble être une observation apparemment contre-intuitive.
Une autre énigme demeure. Pourquoi certaines régions du globe ont plus d’espèces que d’autres ? C’est en particulier le cas des régions équatoriales qui ont un nombre d’espèces plus élevé que les régions polaires. Des dizaines d’hypothèses ont ainsi été formulées, mais aucune ne fait l’objet d’un consensus au sein de la communauté scientifique internationale. Une nouvelle théorie suggère que les mathématiques pourraient jouer un rôle fondamental.
Pour progresser sur ces questions biologiques, la communauté scientifique continue de prélever des êtres vivants (archées, bactéries, végétaux, champignons et animaux) pour les inventorier et étudier leur biologie. Les scientifiques estiment qu’entre 9 % (espèces marines) et 14 % (espèces terrestres) des espèces ont été nommées et décrites à ce jour. Les écologues investiguent les interactions multiples de ces espèces avec l’environnement, y compris le climat, mais aussi avec l’ensemble des espèces vivant autour d’elles, un prérequis essentiel pour comprendre leurs distributions spatiales (biogéographie), leurs rythmes de reproduction (phénologie) et leurs fluctuations temporelles depuis les fluctuations saisonnières jusqu’aux changements centenaires et millénaires, voire même les changements survenant aux échelles de temps géologiques (écologie, paléoécologie et bioclimatologie).
Une nouvelle théorie
Une théorie, baptisée METAL (MacroEcological Theory on the Arrangement of Life), a été proposée pour connecter ensemble biogéographie, phénologie, écologie, paléoécologie et bioclimatologie des espèces, mais aussi pour comprendre comment les communautés d’espèces se forment et comment la biodiversité s’organise et s’altère dans l’espace et le temps.
Comprendre l’organisation spatio-temporelle de la biodiversité à grande échelle nécessite de développer des modèles numériques où les connaissances biologiques, environnementales et climatiques sont mises en équation.
Dans le cadre de la théorie METAL, les bases fondamentales du modèle de biodiversité sont simples. Un grand nombre d’espèces fictives est généré. Chaque espèce fictive (on parle de pseudo-espèce) a des préférences physiologiques uniques, lesquelles définissent leur niche écologique, c’est-à-dire leur réponse face aux contraintes climatiques et environnementales. On peut, dans un premier temps envisager une niche simple, considérant uniquement les dimensions (ou variables) température et disponibilité en eau (ici les précipitations).
La température est un facteur primordial contrôlant la physiologie de toutes les espèces vivant sur notre planète et les précipitations constituent une indication sur la disponibilité en eau, une variable tout aussi importante que la température pour les espèces terrestres. Chacune des millions d’espèces fictives créées interagit avec les facteurs climatiques et colonise progressivement le milieu terrestre et marin (en surface et dans le fond des océans). Au cours des simulations, les espèces s’organisent progressivement en communauté et la biodiversité, plus précisément ici le nombre d’espèces dans une région donnée, est reconstituée.
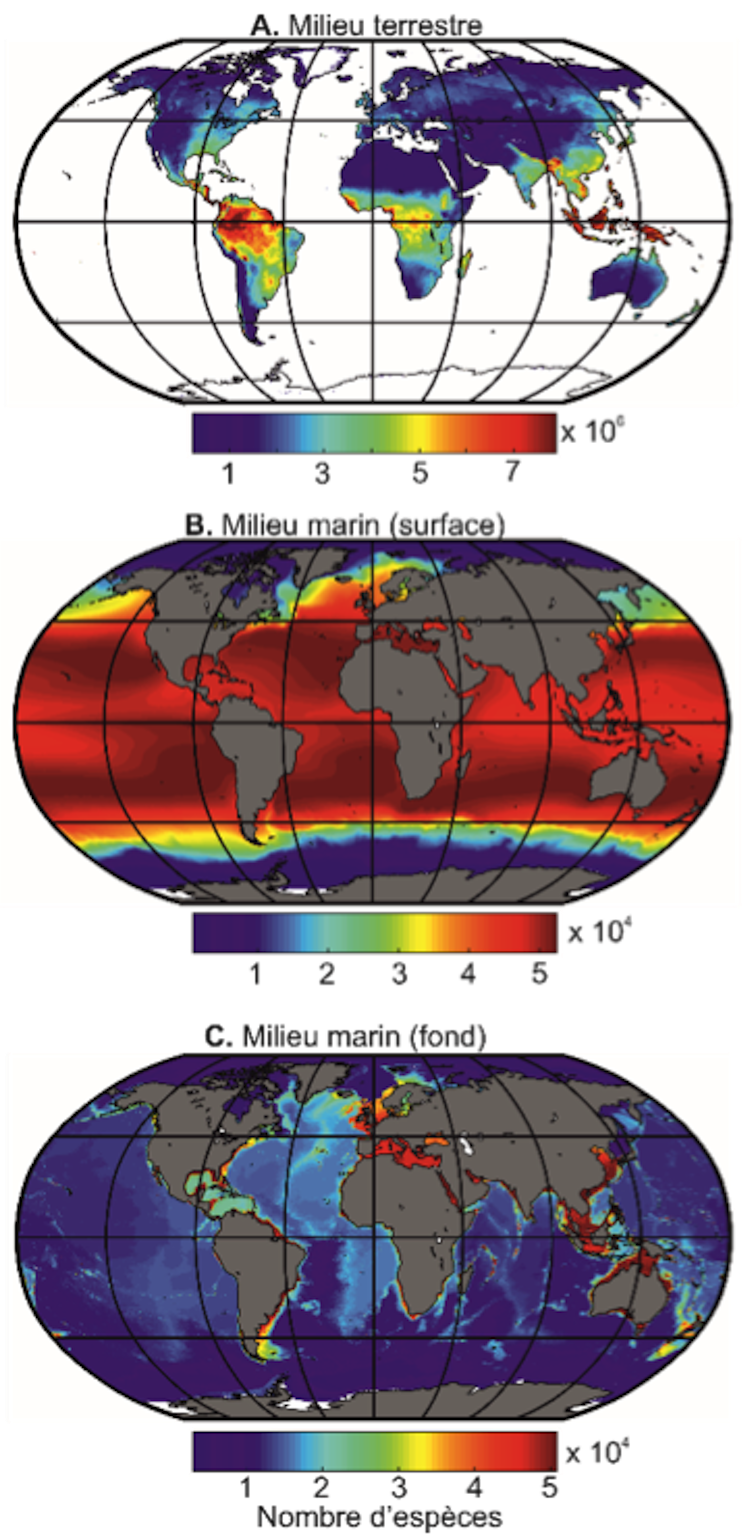
Grégory Beaugrand, Author provided
Ces expériences numériques (ou simulations) reconstituent très correctement la distribution spatiale de la biodiversité telle qu’elle est actuellement observée pour un grand nombre de groupes taxonomiques en milieu terrestre et marin (ex : crustacés, poissons, cétacés, plantes, oiseaux). Les cartes de biodiversité pour les fonds océaniques restent provisoires dans la mesure où peu d’observations ont été réalisées à ce jour pour confirmer ces résultats.
Le grand échiquier de la vie
La reconstitution des distributions de la biodiversité observée dans la nature survient parce que l’interaction niche-climat génère une contrainte mathématique sur le nombre d’espèces pouvant s’établir dans une région donnée. Nous avons nommé cette contrainte le grand échiquier de la vie. Bien qu’il reste une grande part de stochasticité (absence de déterminisme causal) sur le type et le nombre d’espèces pouvant s’établir dans une région, ce nombre ne peut dépasser un seuil théorique fixé par l’interaction niche-climat dans le modèle.
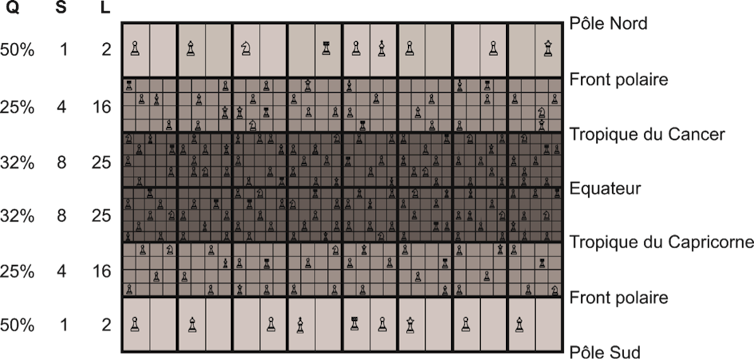
Grégory Beaugrand, Author provided
Par exemple, peu d’espèces peuvent s’établir aux limites minimales et maximales des températures et des précipitations. Les pôles correspondent aux limites minimales des valeurs de températures (pour le milieu terrestre et marin) et de précipitations (pour le milieu terrestre), le nombre d’espèces pouvant s’établir est fondamentalement limité puisque deux espèces ayant la même niche (c’est-à-dire disposant des mêmes adaptations pour faire face aux fluctuations environnementales) ne peuvent coexister d’après le principe d’exclusion compétitive de Gause. Les limites supérieures n’étant pas observées au niveau de l’équateur dans le climat actuel, la biodiversité est maximale au niveau de l’équateur en milieu terrestre et au niveau des régions subtropicales en milieu marin. Ceci n’a pas toujours été le cas et lors de périodes chaudes, la biodiversité pouvait être bien plus basse à l’équateur. De même, la biodiversité a parfois été bien plus haute au niveau des pôles.
Pourquoi le nombre d’espèces terrestres est-il plus important que le nombre d’espèces marines ? Plusieurs facteurs sont à l’origine de cette différence. Le premier, fondamental, est lié aux nombres de dimensions climatiques. L’eau est par définition présente partout dans l’océan ce qui n’est pas le cas pour le milieu terrestre. La dimension climatique supplémentaire augmente fortement le nombre de niches bioclimatiques et ainsi le nombre d’espèces pouvant s’établir dans le milieu terrestre. L’ajout d’une dimension climatique supplémentaire, combiné à des variations géographiques plus prononcées en milieu terrestre, augmente la possibilité de spéciation, c’est-à-dire de création d’espèces, parce qu’un isolement reproductif est plus probable.
Le taux de diversification reste un paramètre important, car il détermine le degré d’occupation des niches dans une cellule géographique donnée et les traits d’histoire de vie de chaque groupe rendent l’échiquier du vivant spécifique à un groupe taxonomique donné ce qui explique parfois le manque d’universalité des patrons de biodiversité à grandes échelles spatiales. L’échiquier de la vie se réorganise en fonction du climat, ce qui le rend dynamique depuis des échelles temporelles fines jusqu’à des échelles très grandes (c.-à-d. les échelles géologiques).
La théorie METAL montre qu’une composante déterministe, c’est-à-dire intelligible et prédictible, contrôle l’organisation des systèmes biologiques depuis le niveau organisationnel de l’individu jusqu’au niveau organisationnel des communautés. Les modèles issus de la théorie METAL, intégrant cette intelligibilité, nous permettent de mieux comprendre l’organisation de la biodiversité sur notre planète, mais aussi de prédire la biodiversité passée contemporaine et future, et donc d’anticiper la réponse des systèmes biologiques face au dérèglement climatique. Les individus sont quant à eux soumis à d’autres règles. Plus les espèces atteignent des tailles importantes, moins elles comportent d’individus, un patron de variabilité connu sous le nom de règle de Damuth (1981).![]()
Grégory Beaugrand, Directeur de recherche CNRS, bioclimatologue, océanologue et écologue, Université de Lille
Cet article est republié à partir de The Conversation sous licence Creative Commons. Lire l’article original.
Ecrire un commentaire